
Heart failure is a global problem with an estimated prevalence of 38 million patients worldwide, a number that is increasing with the ageing of the population. Survival after a diagnosis of heart failure has improved during the past 30 years; however, despite the modest improvement, the 5-year mortality is still approximately 50% – worse than that of many cancers. Efforts to enhance the understanding of the pathobiology of heart failure and to develop new approaches for treatments are direly needed. It is often assumed that the structure of the heart is static. We showed that sarcomeres are highly dynamic structures, and even the largest protein, titin, is replaced in vivo across the entire heart with an 8-day half-life.

The cells in our bodies often live longer than the proteins they contain, so the proteins need to be constantly replaced. However, it is not entirely clear how large protein complexes, composed of multiple different proteins, maintain their structural and functional integrity while replacing their subunits. This is particularly true for cardiomyocytes, which are long-lived cells whose role is to maintain and operate one major protein complex called the sarcomere. As the human heart is estimated to contain more than three trillion sarcomeric units, maintaining sarcomeres must be an energy-intensive process that has to be performed at a very high fidelity. Indeed, studies in human failing hearts using electron microscopy showed significantly reduced sarcomeric myofibrillar density in patients suffering from either idiopathic or ischemic dilated cardiomyopathies, and proteomic studies confirmed the specific loss of sarcomeric proteins in such failing hearts, suggesting proteostasis failure may play an important role in the progression of heart failure.
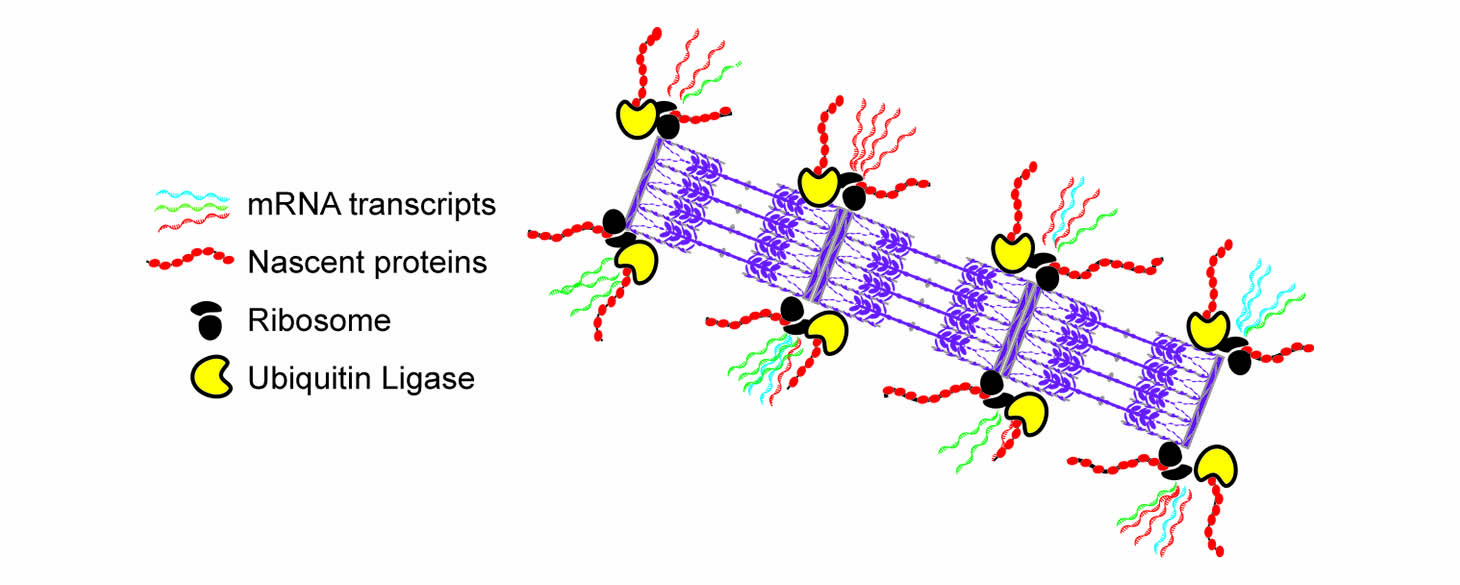
We discovered that sarcomeres contain localized ‘nano-facilities’ positioned on both sides of the Z-line of the sarcomere, where mRNAs, ribosomes, and protein translation are localized. We later identified a ribosomal associated protein called RPSA, and showed that if we knock it out in cardiomyocytes, the cells lose their ribosomal localization, their localized translation, and most of the sarcomeres, showing that localized translation was indeed essential for the maintenance of the sarcomeres. Employing a different labeling scheme of Halo-tagged sarcomeric proteins we imaged and identified the mechanisms underlying the turnover of sarcomeric proteins.
Papers:
Lewis YE, Moskovitz A, Mutlak M, Heineke J, Caspi LH, Kehat I. Localization of transcripts, translation, and degradation for spatiotemporal sarcomere maintenance.
J Mol Cell Cardiol. 2018 Mar;116:16-28.
https://www.ncbi.nlm.nih.gov/pubmed/29371135
Scarborough EA, Uchida K, Vogel M, Erlitzki N, Iyer M, Phyo SA, Bogush A, Kehat I, Prosser BL. Microtubules orchestrate local translation to enable cardiac growth.
Nat Commun. 2021 Mar 11;12(1):1547
https://pubmed.ncbi.nlm.nih.gov/33707436/
Localized translation and sarcomere maintenance requires ribosomal protein SA in mice. Haddad R, Sadeh O, Ziv T, Erlich I, Haimovich-Caspi L, Shemesh A, van der Velden J, Kehat I.
J Clin Invest. 2024 May 14;134(13):e174527
https://pubmed.ncbi.nlm.nih.gov/38743494/
Keeping it fresh: ribosomal protein SA sustains sarcomeric function via localized translation
https://pubmed.ncbi.nlm.nih.gov/38949021/
Imaging of Existing and Newly Translated Proteins Elucidates Mechanisms of Sarcomere Turnover. Douvdevany G, Erlich I, Haimovich-Caspi L, Mashiah T, Prondzynski M, Pricolo MR, Alegre-Cebollada J, Linke WA, Carrier L, Kehat I.
Circ Res. 2024 Jul 4.
https://pubmed.ncbi.nlm.nih.gov/38962864/
Cardiac sarcomere turnover by unidirectional replacement of proteins